Glycoconjugates sit at the crossroads of nearly every major biological recognition event — from how immune cells identify pathogens to how tumors evade detection. These hybrid molecules, formed when carbohydrate chains attach covalently to proteins or lipids, define the biochemical identity of cell surfaces across all living organisms. Understanding their architecture and diversity is a prerequisite for any serious work in glycomics, drug development, or biomarker discovery.
Your three anchors before reading further:
- Glycoconjugates are defined by the covalent bond between a sugar chain (glycan) and a carrier molecule — protein or lipid.
- N-linked and O-linked glycosylation represent the two dominant attachment pathways, each with distinct sequon requirements and cellular machinery.
- The glycome of a single human cell type can express thousands of structurally distinct glycan structures, making it the most information-dense layer of the cell surface.
The field has matured dramatically over the past two decades. Where glycans were once treated as secondary decorations on proteins, glycomics studies now routinely demonstrate that these structures carry functional information governing protein folding, half-life, receptor binding, and immunological tolerance. This guide maps the structural logic of glycoconjugates and their most consequential biological roles.
Researchers entering glycoscience frequently encounter a terminology gap. Terms like monosaccharide, oligosaccharide, glycan, and polysaccharide are used interchangeably in casual speech but carry precise and non-interchangeable definitions in the literature. Clarifying that vocabulary upfront prevents systematic misreading of published protocols and supplier specifications.
Glycans and glycoconjugates: structural definitions
Monosaccharides, oligosaccharides, and polysaccharides
A glycan is the generic term for any carbohydrate chain — whether free or covalently attached to another molecule. The building blocks are monosaccharides: single sugar units such as glucose, galactose, mannose, fucose, N-acetylglucosamine (GlcNAc), N-acetylgalactosamine (GalNAc), and sialic acid. When two to ten of these units link via glycosidic bonds, the resulting structure is called an oligosaccharide. Longer chains crossing that threshold become polysaccharides. The practical boundary matters: most biologically active recognition motifs on cell surfaces are oligosaccharidic (typically 2–15 residues), not fully polymeric.
A glycoconjugate is produced when a glycan chain becomes covalently tethered to a non-sugar carrier. The carrier determines the subclass: if it is a protein, the result is a glycoprotein or proteoglycan; if it is a lipid, the molecule is a glycolipid or lipopolysaccharide. The free glycan and its conjugated form often behave very differently in solution — conjugation alters solubility, conformation, and receptor-binding capacity in ways that isolated oligosaccharides cannot replicate. This distinction has direct consequences for experimental design when selecting reference standards or functional probes for binding assays.
Functionally distinct from simple glycans, the Elicityl research catalog of glycoconjugates spans multiple structural series, including Globo, Isoglobo, blood group antigens, human milk oligosaccharides (HMO), stage-specific embryonic antigens (SSEA), tumor-associated carbohydrate antigens (TACA), and gangliosides, each representing a unique structural family with specialized biological roles.
N-linked vs. O-linked glycosylation
Two major biosynthetic pathways attach glycans to proteins. In N-linked glycosylation, the glycan chain anchors to the amide nitrogen of an asparagine residue, almost exclusively within the canonical sequon Asn-X-Ser/Thr (where X is any amino acid except proline). Assembly occurs co-translationally in the endoplasmic reticulum lumen, beginning with the transfer of a 14-sugar precursor from a dolichol-phosphate carrier. Subsequent trimming and remodeling in the Golgi apparatus generates the three major N-glycan types: high-mannose, complex, and hybrid.
O-linked glycosylation is mechanistically distinct. Sugars attach post-translationally to the hydroxyl groups of serine or threonine residues, with no strict consensus sequon equivalent to the N-glycan Asn-X-Ser/Thr motif. The most widespread form uses GalNAc as the initiating sugar (mucin-type O-glycosylation), but O-fucose, O-GlcNAc, O-mannose, and O-xylose linkages each define separate and functionally specialized pathways. O-GlcNAcylation, for instance, functions almost like a phosphorylation switch on nuclear and cytoplasmic proteins, responding dynamically to metabolic signals.
Sequon and glycan site mapping: Predicting N-glycosylation sites computationally uses the Asn-X-Ser/Thr sequon rule, but not every sequon gets glycosylated — local protein folding and ER accessibility govern the actual occupancy rate. O-glycosylation sites remain harder to predict algorithmically, which is why mass spectrometry-based glycoproteomics workflows remain the most reliable mapping strategy.
The distinction between N- and O-linked pathways carries real experimental consequences. When designing a recombinant glycoprotein for binding studies, the host cell system (CHO, HEK293, insect, yeast) dramatically influences which glycan structures get added and how completely the sites are occupied. The same protein expressed in two different systems can exhibit radically different lectin-binding profiles — a fact the glycomics community has documented repeatedly in comparative expression studies.
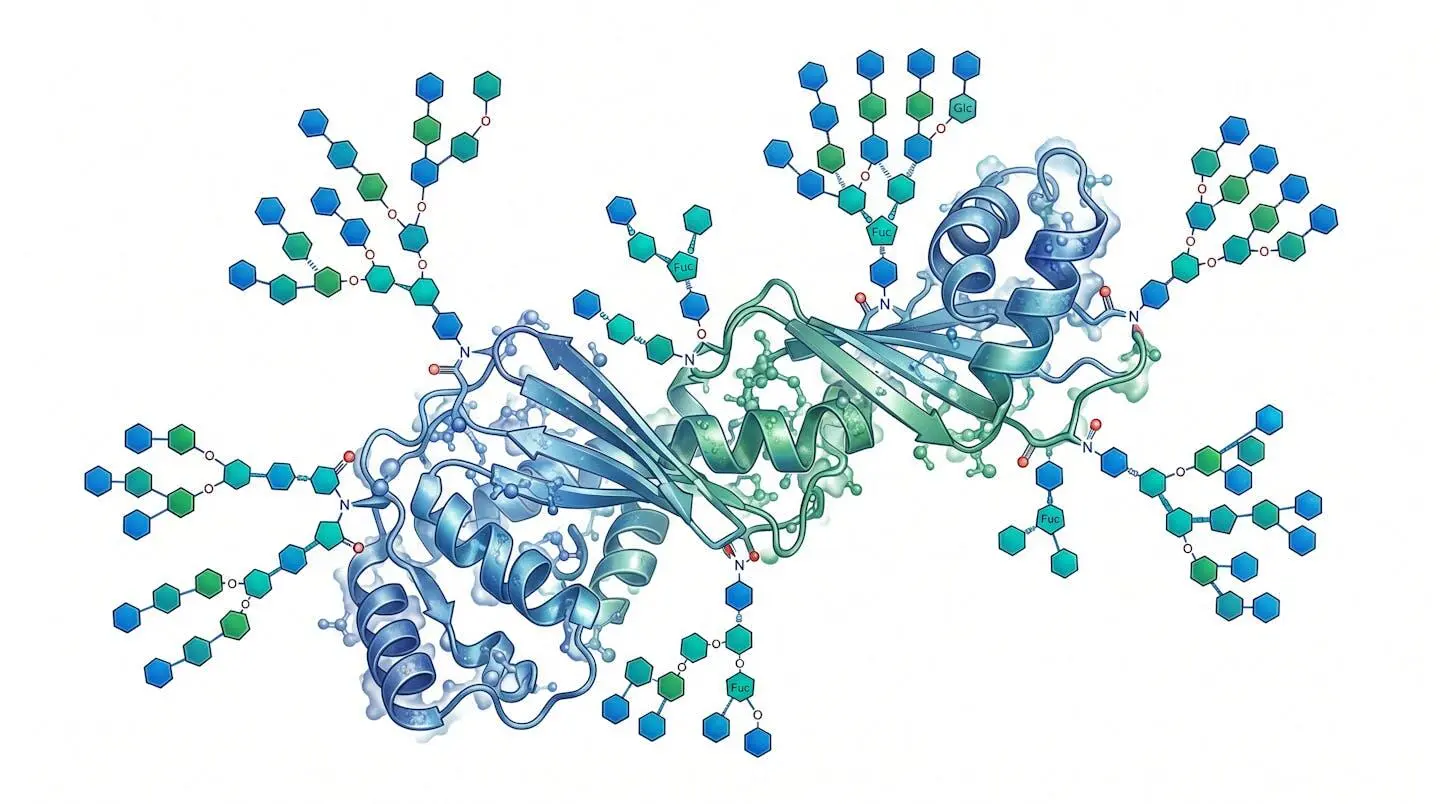
The main classes of glycoconjugates
Glycoproteins and proteoglycans
Glycoproteins are the most abundant glycoconjugate class in mammals. Their sugar content can range from less than 1% by weight to over 80% in mucins — a range that immediately signals the enormous functional diversity within this single category. Most serum proteins, membrane receptors, enzymes, and antibodies are glycoproteins. IgG antibodies, for instance, carry a single complex N-glycan at Asn-297 in the Fc region; subtle changes in that glycan’s terminal residues (sialylated vs. fucosylated vs. afucosylated) shift the antibody’s inflammatory or anti-inflammatory activity measurably, a finding with substantial implications for therapeutic antibody engineering.
Proteoglycans occupy a structurally different niche. Here, the protein core carries long, unbranched glycosaminoglycan (GAG) chains — heparan sulfate, chondroitin sulfate, dermatan sulfate, or keratan sulfate — attached via specific xylose-serine O-linkages. The high negative charge of sulfated GAG chains makes proteoglycans key regulators of extracellular matrix organization, growth factor sequestration, and cell-surface co-receptor function. The precise characterization of amino acid roles at these O-linkage sites remains an active area of investigation, particularly in the context of their involvement in post-translational modification cascades.
Glycolipids and their biological significance
Glycolipids present glycan chains attached to a lipid anchor rather than a protein. The two major subclasses are glyceroglycolipids (common in plant and bacterial membranes) and sphingoglycolipids (dominant in mammalian systems). Within sphingoglycolipids, the ganglioside family deserves specific attention: these are sialic acid-bearing glycosphingolipids concentrated in neural tissue and broadly implicated in neuronal differentiation, synaptic transmission, and several neurodegenerative pathologies.
The Globo and Isoglobo series represent another structurally defined subfamily. Globo-series glycolipids carry a Galα1-4Galβ1-4Glcβ1-Ceramide core, and certain members of this series — notably SSEA-3 and SSEA-4 — serve as stage-specific embryonic antigens marking pluripotent stem cells. This makes them valuable phenotypic markers in cell biology and relevant targets in oncology research, since several tumor-associated carbohydrate antigens (TACA) within the Globo series show elevated expression on cancer cell surfaces compared to normal differentiated tissue.
>50%
Proportion of all mammalian proteins estimated to carry at least one glycosylation site — making glycosylation the most prevalent post-translational modification in the proteome
The structural diversity within glycolipids is generated by the same Golgi-based glycosyltransferase machinery responsible for glycoprotein maturation. This shared biosynthetic infrastructure means that perturbations in glycan metabolism — genetic, pharmacological, or nutritional — tend to affect both glycoprotein and glycolipid populations simultaneously, complicating phenotypic interpretation without careful glycomic profiling.
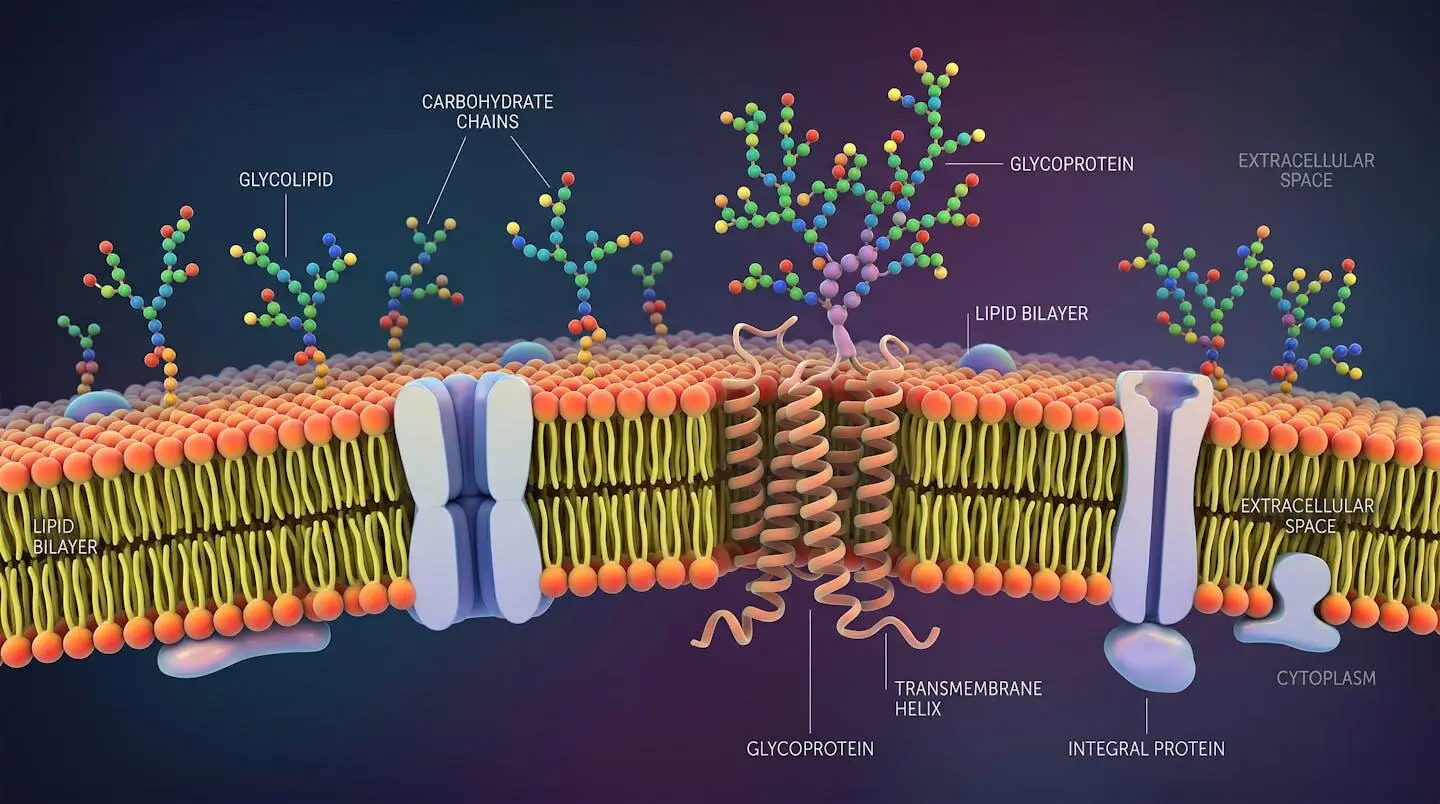
Biological roles of glycosylation in cell signaling and immunity
Glycosylation is not a passive modification. Every major checkpoint in protein quality control within the secretory pathway depends on lectin-mediated reading of glycan structures. Calnexin and calreticulin in the ER retain misfolded glycoproteins specifically through their GlcNAc-trimming state — a classic example of glycan-encoded quality control. Downstream, the sialylation state of surface glycoproteins on hematopoietic cells determines their circulatory lifespan: desialylated glycoproteins expose galactose residues recognized by asialoglycoprotein receptors in the liver, triggering clearance.
In the immune system, the consequences are even more ramifying. Selectin-mediated leukocyte rolling on inflamed endothelium depends on the display of sialyl-Lewis X tetrasaccharide structures on cell-surface glycoproteins. Complement activation, NK cell cytotoxicity regulation through KIR and CD94/NKG2 receptors, and the anti-inflammatory activity of intravenous immunoglobulin preparations all operate through glycan-dependent mechanisms. The glycosylation status of therapeutic antibodies has become a critical quality attribute in biopharmaceutical manufacturing for precisely this reason.
Practical scenario: glycan remodeling in therapeutic antibody production
Consider a typical situation in biopharmaceutical process development. A recombinant IgG1 antibody expressed in CHO cells shows batch-to-batch variability in its afucosylation level — the proportion of Fc glycans lacking core fucose. Afucosylated glycoforms bind FcγRIIIa on NK cells with roughly 50-fold higher affinity than their fucosylated counterparts, translating into substantially enhanced antibody-dependent cellular cytotoxicity (ADCC). When upstream cell culture conditions shift (pH, dissolved oxygen, media composition), the glycosylation profile shifts with them. The process team responds by implementing online glycan monitoring using a lectin-based microarray panel, identifying the fucosylation trajectory early enough to adjust feeding strategies before the batch drifts out of specification. Without glycomic quality control built into the manufacturing workflow, that batch-level variation would only surface during late-stage analytical testing — with significant timeline and cost consequences.
Beyond immunity, glycosylation governs developmental patterning. The Notch signaling pathway is regulated by O-fucosyltransferases and Fringe-family glycosyltransferases that add or extend O-fucose chains on EGF-like repeats in the Notch extracellular domain. Genetic disruption of this machinery — as in Lunatic Fringe knockout mice — produces severe segmentation defects, demonstrating that glycan-level biochemistry encodes body plan information that cannot be substituted by protein sequence alone.
Claim: Glycosylation is a secondary modification that does not substantially affect protein function.
Reality: Glycan structures regulate protein folding stability, circulatory half-life, receptor affinity, and immune checkpoint engagement. In erythropoietin, for instance, removing N-glycans reduces serum half-life from hours to minutes. In the HIV gp120 envelope protein, an N-glycan shield covering roughly half the protein surface actively impedes neutralizing antibody access. These are not marginal effects.
Analytical tools and research applications in glycomics
Characterizing glycoconjugates demands a multi-method approach, because no single technique captures the full picture. Mass spectrometry — particularly LC-MS/MS and MALDI-TOF — provides the highest resolution for glycan composition and sequence. Capillary electrophoresis and high-performance liquid chromatography with fluorescence detection handle quantitative glycan profiling after enzymatic or chemical release. For interaction studies, surface plasmon resonance and isothermal titration calorimetry measure binding kinetics between glycans and lectins or antibodies.
Lectin microarrays have emerged as a particularly productive platform for rapid glycoprofiling. By printing panels of lectins with defined sugar-binding specificities onto solid surfaces, researchers can fingerprint the glycosylation state of cell populations, exosomes, or purified glycoproteins in a single multiplexed experiment. These solutions de microarrays de lectines are now sufficiently standardized to support comparative studies across cell lines, differentiation states, or disease conditions — a shift from purely discovery-mode glycomics toward more systematic glycoprofiling workflows.
Glycoconjugates functionalized for bioconjugation have expanded the experimental toolkit further. Oligosaccharides grafted onto hydroxylated polyacrylamide (PAA) carriers — with alkyne, amine, azide, DBCO, maleimide, or cyclic dithiol functional groups — allow controlled coupling to detection reagents, surfaces, or drug molecules. This chemical versatility is what makes synthetic glycoconjugates indispensable for microarray construction, flow cytometry staining panels, and glycan-targeted drug delivery research. The functional group dictates the conjugation chemistry: strain-promoted azide-alkyne cycloaddition (SPAAC using DBCO-azide pairs) operates under physiological conditions without copper catalysis, making it preferable for cell-surface labeling applications where metal toxicity is a concern.
- Glycan series specificity: Globo, Isoglobo, blood group, HMO, SSEA, or TACA — each series targets distinct biological questions and requires matching reference standards
- Functional group compatibility: the conjugation chemistry must match downstream assay conditions (aqueous pH, temperature, absence of competing nucleophiles)
- Carrier scaffold: PAA-grafted oligosaccharides provide multivalent display mimicking natural cell-surface density, whereas free oligosaccharides are better suited for solution-phase binding assays
- Analytical documentation: NMR characterization and HPLC purity data for defined glycan structures are non-negotiable for publishing reproducible results
The glycome of a given cell type is not fixed. It shifts with differentiation state, metabolic condition, viral infection, and oncogenic transformation. This plasticity is precisely what makes glycomic profiling informative as a biomarker strategy — and what makes it technically demanding, since measurements must be tied to tightly controlled biological conditions to yield interpretable data.

Your priorities for glycomics work
- Identify which glycan series is relevant to your biological question — TACA panels for oncology, HMO series for microbiome-immune crosstalk, SSEA for stem cell phenotyping
- Determine your attachment chemistry requirements before ordering: DBCO-functionalized PAA conjugates for copper-free click labeling, maleimide conjugates for thiol-reactive surfaces, azide conjugates for metabolic labeling strategies
- Validate your lectin panel specificity against known glycan standards before interpreting glycoprofiling data from biological samples
- Cross-reference glycan structure assignments with published glycomics databases (GlyTouCan, GlycoWorkbench) to ensure nomenclature consistency across the literature
The growing availability of defined synthetic glycoconjugates — with characterized structures and precise functional group placement — has removed much of the reagent uncertainty that historically slowed glycomics research. What once required multi-step custom synthesis can now be sourced as validated reference materials, allowing experimental effort to shift toward biological interpretation rather than reagent production. The structural vocabulary covered here — N- and O-glycosylation pathways, glycoprotein versus glycolipid architecture, Globo and TACA series specificity — provides the conceptual scaffold for that work. Cross-reference your glycan structure assignments with GlyTouCan to ensure nomenclature consistency across the literature.
What is the functional difference between a free glycan and a glycoconjugate?
A free glycan in solution exists as a flexible, rapidly tumbling structure with limited multivalency. Once attached to a protein or lipid carrier, the same carbohydrate sequence gains a fixed spatial orientation, increased local concentration on cell surfaces, and altered conformational dynamics that substantially change how lectins and receptors interact with it. Many biological recognition events require the conjugated form — free oligosaccharides can serve as competitive inhibitors or analytical standards, but they do not replicate the clustered display geometry of native glycoconjugates.
Why do tumor cells display altered glycoconjugate patterns?
Oncogenic transformation disrupts the regulated expression and localization of glycosyltransferases and glycosidases within the Golgi apparatus. The resulting biosynthetic rewiring leads to aberrant glycan structures — truncated O-glycans (Tn and sialyl-Tn antigens), overexpressed Globo-series glycolipids, hypersialylation of surface glycoproteins, and loss of normal blood group antigen expression. These tumor-associated carbohydrate antigens (TACA) represent both diagnostic biomarkers and potential targets for glycan-directed immunotherapy approaches currently under active investigation.
What makes polyacrylamide (PAA) a preferred carrier scaffold for glycan display?
Hydroxylated polyacrylamide presents several practical advantages for glycan conjugation studies. It is hydrophilic and chemically inert, minimizing non-specific binding to proteins and surfaces. Multiple oligosaccharide units can be grafted per polymer chain at defined loading densities, creating a multivalent display that better approximates the clustering geometry of glycans on natural cell surfaces than monovalent conjugates. The backbone also tolerates a wide range of functional group chemistries, allowing the same PAA scaffold to carry alkyne, azide, amine, or DBCO groups depending on the downstream conjugation requirement.